¿Cómo surge la vida multicelular?
Modelización de patrones de Fibonacci en tejidos biológicos para revelar mecanismos subyacentes
Resumen
La capacidad de los organismos para organizar con precisión las células dentro de sus tejidos ha permitido la existencia de la vida multicelular en la Tierra durante cientos de millones de años. Sin embargo, el modo en que se produce esta organización celular sigue siendo un enigma. Los mecanismos responsables podrían estar ocultos en los patrones de Fibonacci, ya que muchos tejidos biológicos poseen estructuras geométricas definidas por las relaciones de recurrencia de Fibonacci.
Los datos cuantitativos precisos que describen estos patrones y sus correspondientes ángulos de divergencia ofrecen una oportunidad única para descubrir los mecanismos subyacentes. En biología, la organización de los tejidos surge inicialmente en nichos de células madre que generan estructuras histológicas circulares. Por ello, se desarrolló un modelo de formación de patrones circulares de Fibonacci estructurados por edades.
Los resultados obtenidos sobre el orden de las divisiones celulares y el crecimiento cíclico de los patrones concuerdan con los patrones biológicos de Fibonacci conocidos. La modelización muestra que el tamaño del nicho de células madre y un modo específico de división celular asimétrica determinan la organización de los tejidos multicelulares. Asimismo, ayuda a explicar cómo las células se organizan para formar partes del cuerpo y, en última instancia, un organismo completo durante el desarrollo. Este mecanismo también tiene una gran importancia para comprender la cicatrización de heridas, la formación de tumores y los defectos congénitos.
1. Introducción
Durante los últimos mil millones de años, la vida multicelular evolucionó a medida que la estructura de los tejidos se volvía cada vez más organizada. Esto plantea una cuestión fundamental: si los organismos multicelulares existen gracias a la organización precisa de las células en sus tejidos, ¿qué están haciendo exactamente esas células para formar estructuras tan ordenadas?
La pregunta de cómo se produce la organización celular en los tejidos resulta aún más compleja cuando se comprende que dicha organización no es un proceso estático. Por el contrario, es extraordinariamente dinámica: la organización permanece inalterada mientras el tejido se renueva continuamente mediante divisiones celulares que ocurren dentro de él.
Además, las reglas biológicas responsables de la organización de los tejidos en los organismos multicelulares probablemente aparecieron muy temprano en la evolución y posteriormente fueron conservadas durante el desarrollo de formas de vida más complejas. De hecho, la importancia de reglas biológicas preexistentes como factores que condicionan las trayectorias evolutivas está bien establecida.
Sin embargo, la evidencia disponible se basa principalmente en características anatómicas y no en características celulares. Por ejemplo, el rasgo anatómico innato de poseer cinco dedos (pentadactilia) aparece en numerosas especies. Más recientemente también se descubrieron evidencias de evolución dependiente de la trayectoria en los ojos de los moluscos.
Aun así, cualquiera que haya estudiado histología vegetal o animal sabe que la organización de los tejidos suele estar altamente conservada entre especies muy diferentes. Por ello, es probable que las reglas fundamentales de organización celular sigan presentes hoy en los tejidos de plantas y animales.
El objetivo de este trabajo es estudiar los patrones tisulares de Fibonacci que aparecen con frecuencia en la naturaleza para identificar las leyes matemáticas que describen las reglas celulares responsables de la organización de los tejidos biológicos.
Una característica clave de esta organización es que muchas estructuras histológicas poseen patrones geométricos descritos por sucesiones de Fibonacci. De hecho, los patrones de Fibonacci están tan extendidos en la naturaleza que es probable que el mecanismo de organización tisular se encuentre oculto en ellos.
En las plantas, los números de Fibonacci aparecen con frecuencia en el número de pétalos de las flores, en las espirales de los girasoles o en las espirales de las piñas. En los animales, estos números se observan fácilmente en patrones presentes en la concha del nautilo, las estrellas de mar, las hidras y las medusas.
Los estudios de estas estructuras geométricas han producido una enorme cantidad de datos cuantitativos precisos que describen tanto los patrones numéricos de Fibonacci como sus ángulos de divergencia asociados. Aunque la filotaxia ha proporcionado importantes avances en la descripción cuantitativa de estos patrones, los mecanismos que explican cómo se generan dentro de los tejidos siguen siendo difíciles de comprender.
Por ello, la modelización matemática de los patrones de Fibonacci podría permitir descubrir las leyes matemáticas que explican las reglas biológicas responsables de la organización de los tejidos en organismos multicelulares.
Para abordar este objetivo, en trabajos previos diseñamos modelos matemáticos basados en reglas de división celular con el fin de investigar si podían explicar la formación de patrones de Fibonacci.
En dichos estudios exploramos cómo la división celular asimétrica podría controlar el crecimiento y la renovación de los tejidos multicelulares y dar lugar a patrones asociados a los números de Fibonacci generalizados o números de Fibonacci p. La sucesión de Fibonacci p viene dada por la relación recursiva:
Fn = Fn−1 + Fn−p−1
Estos números suelen representarse geométricamente. En uno de nuestros estudios desarrollamos un modelo de división celular para la organización tisular basado en la biología de la renovación de los tejidos. Dicho modelo se apoyó en el concepto de maduración celular asociado a la división asimétrica propuesto por Spears y Bicknell-Johnson.
La simulación generó patrones dinámicos complejos en los que la organización celular permanecía constante a pesar de la continua división celular que ocurría dentro del tejido. En otro estudio investigamos patrones de ramificación geométrica basados en sucesiones de Fibonacci p, mostrando cómo la regularidad de las ramificaciones puede surgir en sistemas biológicos.
También observamos que la generación de estructuras ramificadas produce patrones de autosimilitud que aparecen a diferentes escalas y dimensiones. En un tercer trabajo analizamos la organización de recubrimientos geométricos de capítulos florales y sus propiedades, incluido el ángulo de rotación responsable de la formación de patrones espirales.
Los resultados mostraron que diferentes estructuras geométricas presentan distintas eficiencias de recubrimiento y que existen patrones organizativos regulares asociados a diferentes proporciones áureas generalizadas. En conjunto, estos estudios sugieren que simples reglas biológicas relacionadas con las asimetrías espaciales y temporales de la división celular pueden comenzar a explicar cómo se mantiene la organización de los tejidos y los patrones geométricos en los organismos multicelulares.
El presente estudio amplía estos trabajos anteriores con el objetivo de identificar los mecanismos celulares que actúan en el nicho de células madre y en el meristemo para controlar la organización de los tejidos. En consecuencia, se investigaron patrones de sucesiones de Fibonacci que conducen a estructuras cíclicas específicas con el fin de comprobar si coinciden con los patrones de Fibonacci y los ángulos de divergencia conocidos en plantas y animales.
La abundante información disponible sobre la organización precisa de distintos meristemos, junto con las medidas cuantitativas de sus ángulos de divergencia, proporciona una oportunidad excepcional para desarrollar modelos capaces de revelar los mecanismos biológicos subyacentes.
2. Biología del meristemo
En biología, el primer paso en la formación de tejidos organizados tiene lugar en el meristemo o en el nicho de células madre, que constituye el plano de organización a partir del cual se desarrolla la estructura del tejido.
En términos generales, el proceso comienza con una célula madre que, tras una o varias divisiones celulares, produce poblaciones de células hijas que forman estructuras anulares o circulares. En algunas plantas existe una única célula inicial capaz de originar toda la planta, mientras que en otras el desarrollo comienza a partir de un grupo de células iniciales.
Durante las primeras fases del desarrollo aparece un patrón bien definido y preciso, denominado primordio, que surge en el ápice celular inicial. Un primordio es una región circular o disco de tejido que se forma en la primera o segunda capa subdérmica del meristemo apical y presenta una elevada actividad proliferativa.
Los primordios aparecen de forma secuencial mediante un proceso de iniciación cíclico, en el que existe un intervalo temporal bien definido entre la formación de un primordio y el siguiente.
Este patrón se cuantifica mediante el ángulo de divergencia, definido como el menor de los dos ángulos determinados en el centro de una sección transversal del ápice de un brote en crecimiento por dos primordios consecutivos.
La conservación precisa de este ángulo resulta esencial para mantener la estructura y la organización del tejido. Además, el ángulo de divergencia probablemente siga siendo igual de importante en los tejidos adultos, ya que éstos permanecen en continua renovación durante toda la vida del organismo.
Aunque la generación de patrones geométricos derivados del ángulo de divergencia —como las estructuras ramificadas o los capítulos florales— ha sido ampliamente estudiada, la formación de estructuras circulares ha recibido mucha menos atención.
Por esta razón, en este trabajo se desarrolla un modelo de formación de patrones circulares de Fibonacci estructurados por edades, con el objetivo de investigar cómo surgen los patrones de primordios en distintos tejidos a partir de sus ángulos de divergencia conocidos y de las sucesiones de Fibonacci asociadas.
3. Enfoque de modelización
El modelo de formación de círculos de Fibonacci estructurados por edades se basa en la combinación de dos propuestas publicadas previamente:
- Las reglas de división celular asimétrica propuestas por A. P. Berdyshev.
- El modelo de formación de patrones circulares y crecimiento circunferencial inspirado en la denominada Ley de Parkinson.
Las reglas de división celular asimétrica fueron publicadas en 1972 por el matemático ruso A. P. Berdyshev. Su modelo mostraba que los números de Fibonacci podían generarse mediante una estructura basada en la edad de las células, donde éstas nacen, maduran, se reproducen y finalmente mueren siguiendo un conjunto de reglas para la división celular asimétrica.
Este modelo producía exactamente la misma sucesión numérica que el célebre problema de los conejos planteado originalmente por Fibonacci, con la diferencia de que incorporaba una estructura de edades mucho más parecida a la organización real de los tejidos multicelulares.
Ese mismo año apareció otro modelo singular publicado por J. M. Holland en su libro Studies in Structure. Holland propuso un mecanismo de formación de patrones circulares y crecimiento periférico inspirado en la expansión de los comités burocráticos descrita por la Ley de Parkinson.
Este modelo resulta especialmente interesante porque genera patrones circulares en crecimiento y muchos tejidos biológicos poseen precisamente una estructura tubular o epitelial. La formación de estos tubos requiere un proceso dinámico de división celular que produce configuraciones cíclicas.
Con el fin de comprender mejor los mecanismos celulares responsables de la organización de los tejidos, el modelo desarrollado en este trabajo combina el concepto de estructura por edades de Berdyshev con las ideas de Holland sobre la formación de patrones circulares.
4. Diseño del modelo
Se desarrolló un modelo dinámico de formación de círculos de Fibonacci estructurados por edades que simula las primeras etapas del desarrollo de un tejido. El proceso comienza en el nicho de células madre, donde una o varias células iniciales generan, mediante sucesivas divisiones, poblaciones celulares que forman estructuras anulares o circulares.
Las reglas del modelo incorporan la edad de cada célula, el número de células iniciadoras, la dirección alternante de las divisiones, el orden en que éstas se producen y el número concreto de divisiones que experimentan las células descendientes.
Las reglas fundamentales para la proliferación y maduración celular son las siguientes:
- Nacimiento celular. Una o varias células iniciadoras permanecen en el nicho de células madre. Cuando una de ellas se divide, la célula hija más joven permanece en el nicho mientras que la más antigua inicia su proceso de maduración. El número de células iniciadoras viene determinado por la cantidad de células existentes antes de que, en un mismo intervalo temporal, se produzcan múltiples divisiones celulares. De este modo, el sistema puede interpretarse como formado por dos zonas de crecimiento: el nicho de células madre y una zona proliferativa responsable de generar el patrón de Fibonacci.
- Maduración celular. Las células hijas deben atravesar un periodo de maduración antes de poder dividirse. En concreto, cada célula permanece inactiva durante un número determinado de generaciones antes de iniciar nuevas divisiones.
- División celular. Una vez maduras, las células se dividen siguiendo un número prefijado de divisiones, un orden determinado y una dirección específica. Estas divisiones tienen lugar con una frecuencia constante.
- Muerte celular. Tras completar un número determinado de divisiones, las células dejan de dividirse y mueren, aunque permanecen formando parte de la estructura circular de Fibonacci.
- Estructura circular de Fibonacci. Aplicando las reglas anteriores, la secuencia de divisiones celulares genera siempre una disposición circular perfectamente definida. La célula o células iniciales del nicho de células madre dan origen a toda la estructura circular.
5. Resultados
El modelo basado en una estructura de edades generó dos tipos principales de resultados:
- el orden en que se producen las divisiones celulares;
- los patrones circulares obtenidos, que posteriormente se compararon con las sucesiones de Fibonacci y los ángulos de divergencia observados en los tejidos biológicos.
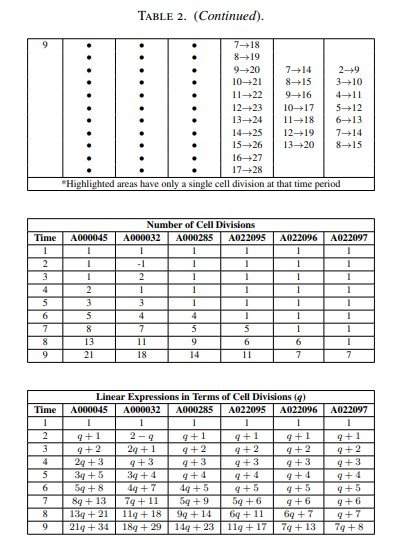
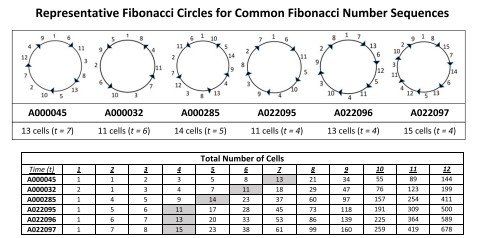
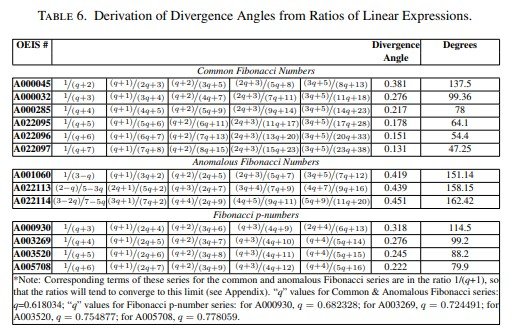
La Tabla 1 recoge, utilizando la numeración de la On-Line Encyclopedia of Integer Sequences (OEIS), las sucesiones de Fibonacci más comunes, varias sucesiones anómalas y las sucesiones generalizadas de Fibonacci p, junto con sus correspondientes ángulos de divergencia.
Para comprobar si el modelo reproducía estos patrones se siguió un proceso en dos etapas. En primer lugar, utilizando las reglas de división celular dependientes de la edad propuestas por Berdyshev, se estudió si el orden de las divisiones celulares generaba una sucesión de generaciones sucesivas que coincidiera con cada una de las sucesiones de Fibonacci. Posteriormente se analizó si esos resultados permitían deducir el ángulo de divergencia, como se explica al final de esta sección.
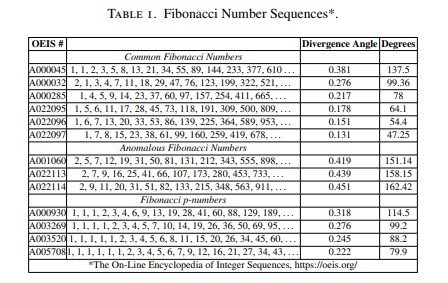
Modelización de las sucesiones de Fibonacci comunes
En primer lugar se determinó la estructura basada en la edad correspondiente a las sucesiones de Fibonacci más comunes con el objetivo de establecer el orden de las divisiones celulares.
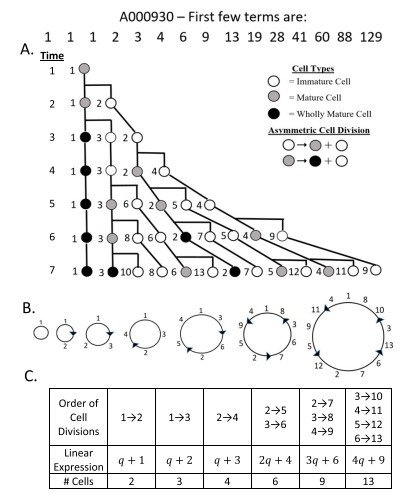
Como ejemplo, la Figura 1A muestra el árbol de divisiones celulares que produce, con el paso del tiempo, una secuencia que coincide con la sucesión generalizada de Fibonacci A000045.
Una vez establecida esta estructura basada en la edad, fue posible construir el correspondiente patrón circular de Fibonacci (Figura 1B) siguiendo el orden de las divisiones celulares mostrado en la Figura 1C.
También se obtuvo una expresión lineal para las distintas edades celulares, es decir, para el número de células nuevas y el número de células procedentes de generaciones anteriores presentes en cada instante temporal. Obsérvese que estas progresiones geométricas lineales también pueden expresarse como una función del número de células nuevas generadas (q), tal como se explica en el Apéndice C.
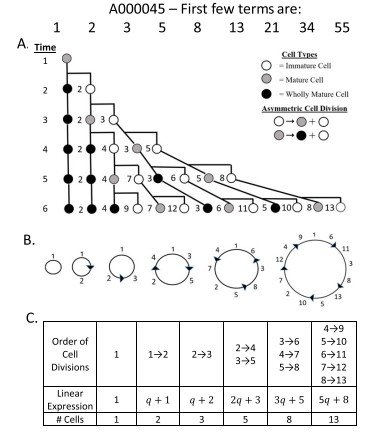
De manera análoga (véase la Figura 2), se modelizó el orden de las divisiones celulares para determinar la sucesión de generaciones que reproduce la sucesión de Lucas (A000032).
Una vez establecido este orden de divisiones, se obtuvo el correspondiente patrón circular de Fibonacci y las expresiones lineales asociadas a las distintas edades celulares para la sucesión de Lucas. Posteriormente, el mismo procedimiento se aplicó al resto de las sucesiones comunes de Fibonacci (Tabla 2).
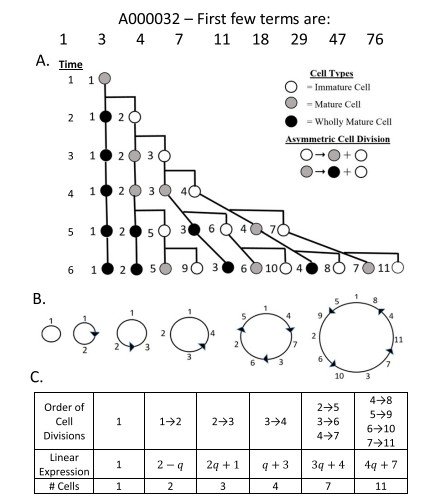
El análisis de todas las sucesiones comunes de Fibonacci reveló un patrón muy interesante que aparece de forma sistemática en todas ellas. En concreto, se observa un incremento progresivo del número de células iniciadoras presentes antes de que, durante un mismo intervalo temporal, tengan lugar múltiples divisiones celulares (más de dos). En la Tabla 2 estas situaciones aparecen resaltadas.
La modelización mediante árboles de divisiones celulares, siguiendo las reglas de Berdyshev, permite visualizar claramente este comportamiento, como se muestra en la Figura 3.
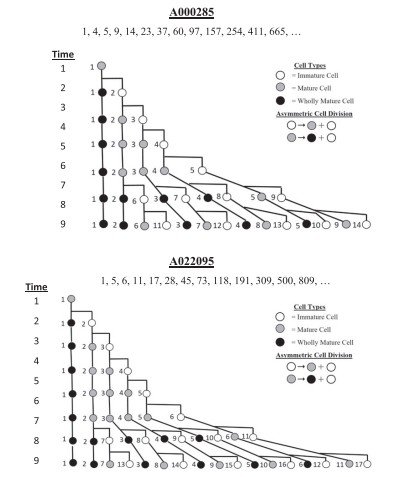
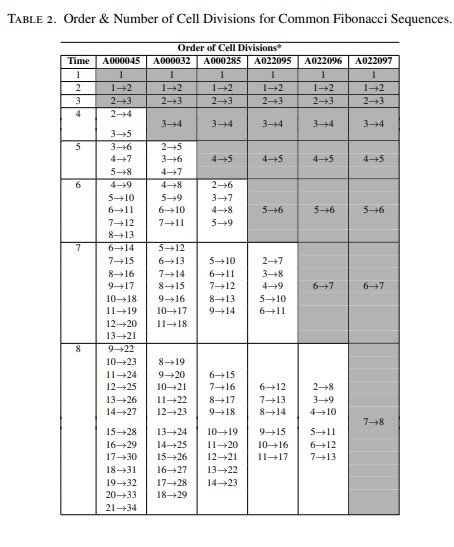
En particular, el patrón de generaciones sucesivas correspondiente a las sucesiones comunes de Fibonacci A000285 y A022095 revela la existencia de un mayor número de células iniciadoras (cinco y seis, respectivamente) en comparación con las sucesiones A000045 y A000032, que requieren únicamente tres y cuatro células iniciadoras (véanse las Figuras 1 y 2).
Las restantes sucesiones comunes de Fibonacci (A022096 y A022097) presentan un número aún mayor de células iniciadoras, tal como puede observarse en la Tabla 2.
Este resultado indica que los distintos meristemos poseen un número diferente de células iniciadoras en su nicho de células madre. En consecuencia, el tamaño del nicho de células madre —es decir, el número de células iniciadoras— constituye un factor determinante en la organización celular del meristemo.
De hecho, para cada una de las sucesiones comunes de Fibonacci estudiadas se obtuvo un patrón circular de Fibonacci específico, como se mostrará en la Figura 4.
Modelización de las sucesiones anómalas de Fibonacci
También se determinó la estructura basada en la edad para las sucesiones anómalas de Fibonacci con el fin de establecer el orden de las divisiones celulares.
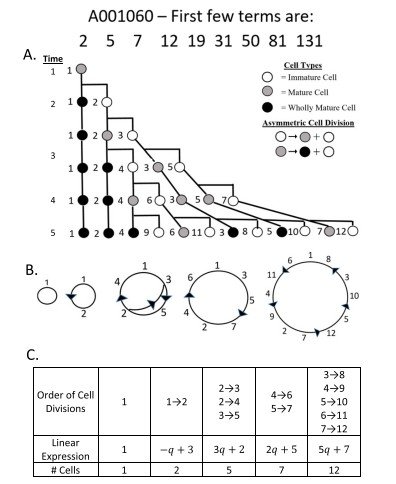
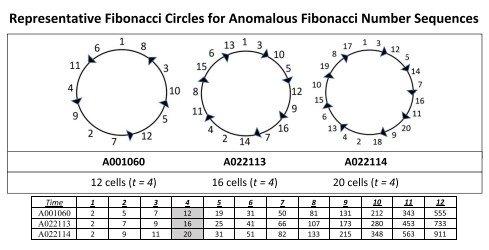
Como ejemplo, la Figura 5 muestra el árbol de divisiones correspondiente a la sucesión de generaciones que reproduce la sucesión anómala A001060, así como el patrón circular de Fibonacci obtenido a partir del orden de dichas divisiones.
La Figura 6 presentará los patrones circulares deducidos para las tres sucesiones anómalas de Fibonacci: A001060, A022113 y A022114.
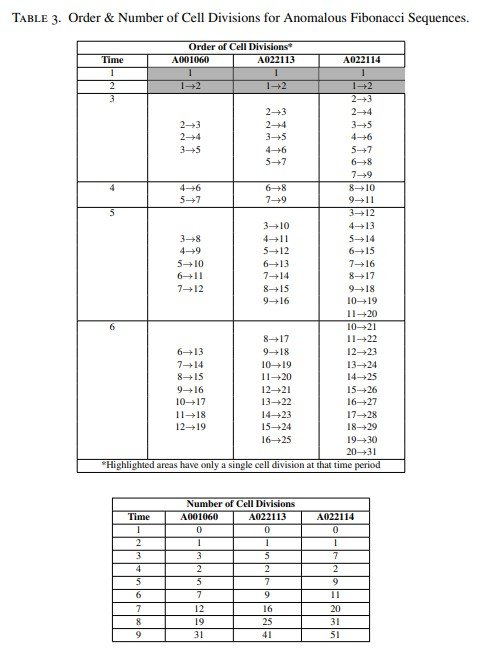
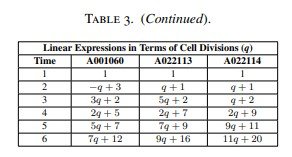
El orden de las divisiones celulares y las expresiones lineales correspondientes a estas sucesiones se recogen en la Tabla 3.
Debe observarse que, en los tres casos, el número de células iniciadoras es siempre de dos células. La diferencia entre las estructuras basadas en la edad de estas tres sucesiones anómalas reside en el número variable de divisiones celulares que tienen lugar durante el tercer ciclo temporal tras la formación del nicho de células madre.
En otras palabras, esta diferencia aparece porque una célula madre y una o varias de sus células hijas pueden dividirse más de una vez durante ese tercer ciclo, tal como muestran las Figuras 5 y 6 y la Tabla 3.
Modelización de las sucesiones de Fibonacci p
También se determinó la estructura basada en la edad correspondiente a las sucesiones generalizadas de Fibonacci p. Como ejemplo, la Figura 7 muestra el árbol de divisiones celulares que reproduce la sucesión A000930, así como el patrón circular de Fibonacci obtenido a partir del orden de dichas divisiones.
Un aspecto especialmente relevante es que aparece un retraso adicional de un ciclo temporal en la maduración de las células inmaduras antes de que puedan convertirse en células maduras.
Asimismo, las reglas que determinan la dirección de las divisiones celulares y el número concreto de divisiones experimentadas por las células hijas también varían entre las distintas sucesiones de Fibonacci p.
Los patrones circulares específicos deducidos para estas sucesiones se presentarán en la Figura 8, mientras que el orden de las divisiones celulares y las expresiones lineales correspondientes se recogen en la Tabla 4.
Debe señalarse, además, que el número de células iniciadoras aumenta progresivamente desde 4 hasta 7 células en las sucesiones de Fibonacci p, tal como se indica en las zonas sombreadas de la Tabla 4.
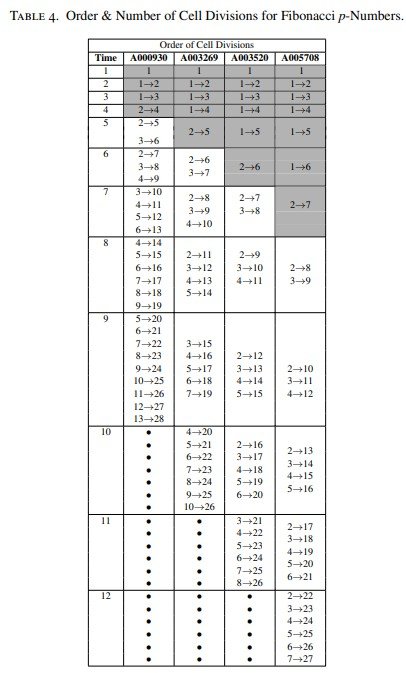
Modelización de los ángulos de divergencia
Una vez identificadas las estructuras basadas en la edad que reproducen cada una de las sucesiones de números de Fibonacci estudiadas, se investigó si los resultados obtenidos por el modelo permitían deducir el correspondiente ángulo de divergencia.
Los resultados muestran que estos ángulos pueden obtenerse tanto a partir del número de células presentes en ciclos temporales sucesivos (Tabla 5) como mediante los cocientes entre las expresiones lineales que describen el crecimiento celular (Tabla 6).
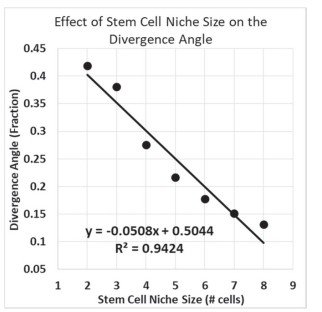
Además, el estudio revela la existencia de una relación lineal entre el tamaño del nicho de células madre, expresado como el número de células iniciadoras, y el ángulo de divergencia (Figura 9).
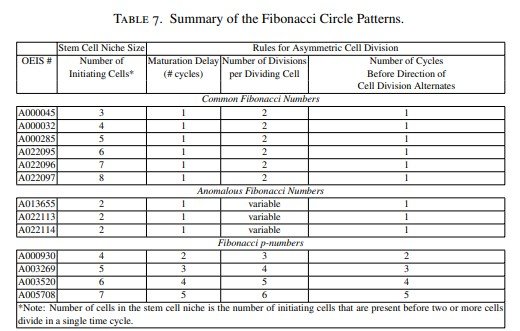
Finalmente, las diferencias observadas entre los distintos patrones de Fibonacci en cuanto al tamaño del nicho de células madre y a las reglas de división celular se resumen en la Tabla 7.
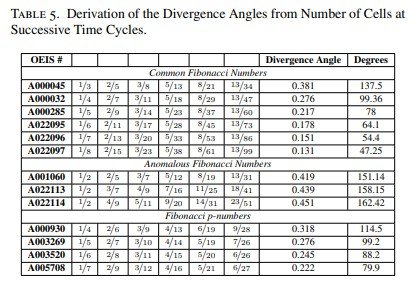
6. Discusión
La modelización de patrones circulares de Fibonacci estructurados por edades demuestra que un conjunto bien definido de reglas celulares es capaz de producir los patrones de números de Fibonacci conocidos en la naturaleza, junto con sus correspondientes ángulos de divergencia.
En efecto, los patrones obtenidos mediante el modelo reproducen las sucesiones comunes, las sucesiones anómalas y las sucesiones de Fibonacci p, así como sus respectivos ángulos de divergencia. Estas variaciones se obtuvieron introduciendo modificaciones en las reglas del modelo relacionadas con el número de células iniciadoras, el número de divisiones celulares, el orden en que dichas divisiones tienen lugar, la dirección de la división celular y el retraso en la maduración de las células.
La incorporación de estas reglas permitió generar un patrón circular de Fibonacci característico para cada una de las sucesiones estudiadas y para cada uno de sus ángulos de divergencia. En particular, el número de células que aparecen sucesivamente en un instante determinado coincide con las distintas sucesiones de Fibonacci conocidas.
Además, el crecimiento de los círculos de Fibonacci se produce por expansión a lo largo de toda la circunferencia y no desde un único punto central. Este crecimiento circunferencial concuerda con la forma en que muchas estructuras circulares se desarrollan en la naturaleza.
El modelo también resulta coherente con el proceso continuo de renovación de los tejidos, en el que las divisiones celulares tienen lugar de manera permanente mientras la organización global del tejido permanece estable.
Los resultados muestran asimismo que el tamaño del nicho de células madre, determinado por el número de células iniciadoras, influye directamente en la organización de los patrones de Fibonacci.
Una vez fijado el tamaño del nicho de células madre, el modelo predice una estructura bien definida en la que el crecimiento del círculo sigue las reglas descritas por la ley de Parkinson. De hecho, la simulación demuestra que pueden generarse patrones circulares específicos asociados a distintos tamaños del nicho de células madre, incluso manteniendo constantes las demás reglas de división celular.
Además, se observa una relación lineal entre el tamaño del nicho de células madre y el valor del ángulo de divergencia, resultado que constituye uno de los hallazgos más relevantes del modelo desarrollado.
Sin embargo, el tamaño del nicho de células madre no es el único factor que afecta a la estructura de los patrones circulares generados.
En efecto, el estudio de las sucesiones anómalas de Fibonacci muestra que, aunque el número de células iniciadoras permanece constante, las reglas de división celular cambian durante el tercer ciclo temporal tras la formación del nicho de células madre.
Este resultado pone de manifiesto que pequeñas modificaciones en las reglas de división celular pueden producir diferencias apreciables en la organización de los tejidos. En otras palabras, variaciones en el modo en que se divide una célula pueden traducirse en patrones tisulares diferentes.
En las sucesiones de Fibonacci p los cambios afectan tanto al tamaño del nicho de células madre como al modo de división celular. El número de células iniciadoras aumenta a medida que se consideran distintas estructuras basadas en la edad para estas sucesiones.
Asimismo, las reglas de división celular difieren en varios aspectos: el retraso de maduración antes de que las células inmaduras puedan dividirse, el número de divisiones que experimenta cada célula y el número de ciclos que transcurren antes de que cambie la dirección de la división celular.
Estas diferencias en el tamaño del nicho de células madre y en las reglas de división afectan al ritmo de crecimiento de los círculos de Fibonacci, produciendo números de células distintos en cada instante temporal y dando lugar a las diferentes sucesiones de Fibonacci p.
Una vez determinadas las estructuras basadas en la edad correspondientes a cada patrón de Fibonacci, el modelo demuestra que el ángulo de divergencia puede obtenerse a partir del número de células presentes en ciclos temporales sucesivos y también mediante los cocientes entre las expresiones lineales del modelo.
En conjunto, estos resultados indican que tanto el modo específico de división celular como el tamaño del nicho de células madre determinan el valor del ángulo de divergencia.
7. Resumen y conclusiones
La modelización de la formación de patrones circulares de Fibonacci estructurados por edades indica que un conjunto específico de reglas biológicas puede dar lugar a las diferentes estructuras de Fibonacci observadas en la naturaleza.
En primer lugar, el modelo reproduce correctamente la sucesión generalizada de Fibonacci A000045 y genera un círculo de Fibonacci con un ángulo de divergencia de 137,5°, valor característico de numerosos patrones naturales.
Además, el modelo reproduce también las restantes sucesiones comunes, las sucesiones anómalas y las sucesiones de Fibonacci p, junto con sus respectivos ángulos de divergencia.
Las diferencias entre estos patrones aparecen como consecuencia de variaciones en el tamaño del nicho de células madre y en las reglas que gobiernan la división celular asimétrica, tal como se resume en la Tabla 7.
Estos patrones circulares de Fibonacci estructurados por edades muestran que un conjunto relativamente sencillo de reglas es suficiente para controlar la formación de la estructura de los tejidos tanto en plantas como en animales.
En consecuencia, el estudio de estos patrones revela que los mecanismos biológicos responsables de la organización de los tejidos dependen de reglas relacionadas con la edad de las células, su proceso de división y su maduración.
La principal aportación de este trabajo es mostrar que determinadas reglas biológicas relativas a la división celular asimétrica y al tamaño del nicho de células madre son factores clave para explicar la organización de los tejidos en los organismos vivos.
Este hallazgo no solo mejora nuestra comprensión de cómo se forman y mantienen los tejidos durante el desarrollo, sino que también puede contribuir a explicar procesos biológicos tan importantes como la cicatrización de heridas, el desarrollo del cáncer y la aparición de determinados defectos congénitos, todos ellos estrechamente relacionados con la organización de los tejidos.
Apéndices
Apéndice A. Expresión geométrica de los números de Fibonacci p
Los números de Fibonacci p suelen representarse mediante una interpretación geométrica basada en la razón áurea generalizada.
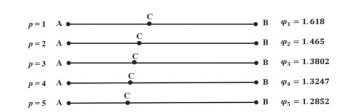
La razón áurea p se define a partir de las cantidades CB y AC cuando su razón es igual a la razón entre las dos cantidades mayores AB y CB, elevada a la potencia p. Es decir, si
\[ AB>CB>AC, \]
entonces se verifica la relación
\[ \varphi_p = \frac{CB}{AC} = \left( \frac{AB}{CB} \right)^p. \]
donde la letra griega \(\varphi_p\) representa la razón áurea generalizada de orden p. Esta cantidad es la raíz positiva de la ecuación
\[ \varphi_p^{\,p+1}-\varphi_p^{\,p}-1=0. \]
Los números de Fibonacci p, denotados por \(F_p\), constituyen una generalización natural de los números de Fibonacci clásicos. La sucesión de Fibonacci p satisface la relación de recurrencia
\[ F_n = F_{n-1} + F_{n-p-1}. \]
Cuando \(p=1\) se recupera la sucesión clásica de Fibonacci. Para valores mayores de \(p\), el término siguiente depende del término inmediatamente anterior y del situado \(p+1\) posiciones antes, dando lugar a nuevas familias de sucesiones con propiedades geométricas análogas.
La sucesión de Fibonacci p se define mediante la relación de recurrencia
\[ F_p(n)=F_p(n-1)+F_p(n-p-1), \qquad n>p+1, \]
con las condiciones iniciales
\[ F_p(0)=0, \qquad F_p(1)=F_p(2)=\cdots=F_p(p)=F_p(p+1)=1. \]
Los números de Fibonacci p están estrechamente relacionados con la razón áurea generalizada. Cuando \(p=1\) se obtiene la sucesión clásica de Fibonacci y la razón áurea habitual. En consecuencia, los números de Fibonacci clásicos están relacionados con la razón áurea clásica.
La conocida fórmula de Binet expresa el término \(n\)-ésimo de la sucesión de Fibonacci en función de \(n\) y de la razón áurea. Cuando \(n\) aumenta, el cociente entre dos términos consecutivos converge hacia dicha razón:
\[ F_n= \frac{\varphi^n-\psi^n}{\varphi-\psi} = \frac{\varphi^n-\psi^n}{\sqrt5}, \]
\[ \varphi=\frac{1+\sqrt5}{2}\approx1.618034, \qquad \psi=\frac{1-\sqrt5}{2}\approx-0.618034. \]
Kilic y Stakhov obtuvieron posteriormente una generalización de la fórmula de Binet para las sucesiones de Fibonacci p.
Apéndice B. La ley de Parkinson
La Ley de Parkinson, aplicada al crecimiento del personal y a la expansión de los comités, establece las siguientes reglas:
- El primer miembro permanecerá un año en el comité antes de convertirse en miembro asesor. Todos los demás miembros actuarán como miembros con derecho a voto durante exactamente dos años y, posteriormente, pasarán a ser miembros asesores o abandonarán el comité.
- Durante cada año de servicio, cada miembro con derecho a voto propondrá la incorporación de un nuevo miembro al comité.
- Cada miembro recibe un número de rango según su antigüedad. El rango 1 se asigna al primer miembro, los rangos 2 y 3 a los siguientes, y así sucesivamente. Cada miembro conserva su rango incluso después de retirarse.
- Los nuevos miembros son incorporados respetando el orden de antigüedad de quienes los proponen.
- En los años pares, los nuevos miembros ocupan su lugar a la izquierda del miembro que los propone en la mesa del comité; en los años impares lo hacen a su derecha.
- Los miembros asesores continúan asistiendo a las reuniones del comité de planificación. Si no pueden hacerlo debido a otras obligaciones, pasan automáticamente a la categoría de miembros retirados.
Apéndice C. Expansión de las progresiones geométricas como función de \((q+1)\)
Se ha observado que el recíproco de la razón áurea generalizada, denominado cociente \(q\), desempeña un papel fundamental en las sucesiones de Fibonacci p. Se define como
\[ q=\frac1{\varphi_p}. \]
A partir de la ecuación característica
\[ \varphi_p^{\,p+1}-\varphi_p^{\,p}=1, \]
y sustituyendo \(q=1/\varphi_p\), se obtiene
\[ q=1+q^p, \]
de donde también se deduce
\[ q^{-1}=1+q^{\,p-1}, \]
lo que conduce a
\[ 1=q+q^{\,p+1}. \]
En el caso particular \(p=1\), esta expresión puede escribirse como
\[ \frac1{1-q} = \frac1q, \]
de donde se obtiene
\[ q^2=1-q. \]
A partir de esta identidad es posible describir el crecimiento de los círculos de Fibonacci mediante progresiones geométricas expresadas como función de \((q+1)\).
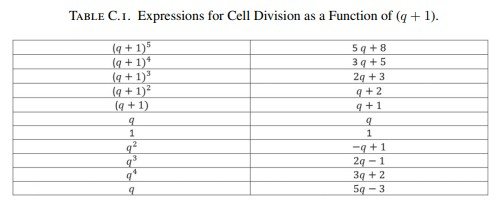
Por ejemplo,
\[ (q+1)^2=q^2+2q+1. \]
Sustituyendo \(q^2=1-q\),
\[ (q+1)^2=q+2. \]
De forma análoga,
\[ (q+1)^3=(q+1)(q+1)^2, \]
y sustituyendo nuevamente la expresión obtenida para \((q+1)^2\),
\[ (q+1)^3=2q+3. \]
Repitiendo este procedimiento de manera sucesiva pueden obtenerse todas las expresiones de la progresión geométrica como funciones de las distintas potencias de \((q+1)\).
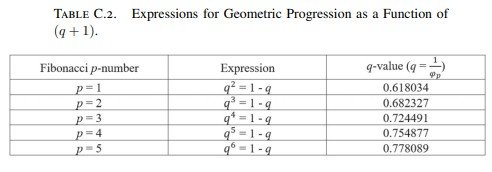
Para las sucesiones de Fibonacci p, la razón correspondiente viene dada por
\[ q=\frac1{\varphi_p} = \left(\frac1{\varphi}\right)^n, \]
de modo que
\[ q^{\,n+1}=1-q, \]
donde \(n=p\). Por ejemplo, cuando \(p=2\),
\[ q^3=1-q. \]
Agradecimientos
El autor expresa su agradecimiento al Dr. Gilberto Schleiniger y a los miembros del equipo de investigación del Center for Applications of Mathematics in Medicine, de la Universidad de Delaware, por su apoyo y por las valiosas discusiones mantenidas durante el desarrollo de este trabajo.
Asimismo, agradece especialmente al Dr. Nicholas Petrelli y al Cawley Center for Translational Cancer Research por su apoyo al desarrollo de modelos matemáticos aplicados a la investigación biomédica.
Declaración de conflictos de interés
El autor declara que no existe ningún conflicto de intereses relacionado con este trabajo.
Financiación
Este trabajo fue financiado parcialmente gracias al generoso apoyo de la Lisa Dean Moseley Foundation for Stem Cell Research, BioCode Innovations Inc. y CATX Inc.
ORCID
Bruce M. Boman
https://orcid.org/0000-0002-1335-9149
Referencias
Las referencias bibliográficas se mantienen en su idioma original para facilitar su localización y consulta.
Clasificación MSC 2020: 11B39, 33C05
Centro de Aplicaciones de las Matemáticas en Medicina
Departamento de Ciencias Matemáticas
Universidad de Delaware
Newark, Delaware 19716, EE. UU.
Correo electrónico: BrBoman@udel.edu
Este artículo ha sido republicado con pequeñas modificaciones que no afectan a su contenido académico.
Enlaces relacionados
Si deseas profundizar en la filotaxis, las fracciones continuas y el ángulo de divergencia, puedes consultar las siguientes páginas.